Tartrate-resistant acid phosphatase
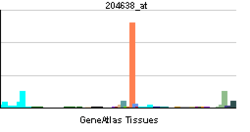
| View/Edit Human | View/Edit Mouse |
Tartrate-resistant acid phosphatase (TRAP or TRAPase), also called acid phosphatase 5, tartrate resistant (ACP5), is a glycosylated monomeric metalloprotein enzyme expressed in mammals.[3] It has a molecular weight of approximately 35kDa, a basic isoelectric point (7.6-9.5), and optimal activity in acidic conditions. TRAP is synthesized as latent proenzyme and activated by proteolytic cleavage and reduction.[4][5] It is differentiated from other mammalian acid phosphatases by its resistance to inhibition by tartrate and molecular weight.
The mechanism of phosphate ester hydrolysis by TRAP is through a nucleophilic attack mechanism,[6] whereby, catalysis occurs with the binding of a phosphate-substrate to the Fe2+ in the active site of TRAP. This is then followed by a nucleophilic attack by a hydroxide ligand on the bound phosphorus atom, resulting in cleavage of the phosphate ester bond and production of an alcohol. The exact identity and mechanism of the hydroxide ligand is unclear, but it is thought to be either a hydroxide that bridges the metal ions within the active site or a terminal hydroxide bound to Fe3+, with conflicting reports for both mechanisms.
TRAP expression and cell localization
Under normal circumstances, TRAP is highly expressed by osteoclasts, activated macrophages, neurons, and by the porcine endometrium during pregnancy.[7][8] In newborn rats, TRAP is also detectable in the spleen, thymus, liver, kidneys, skin, lung, and heart at low levels. TRAP expression is increased in certain pathological conditions. These include leukaemic reticuloendotheliosis (hairy cell leukaemia), Gaucher’s disease, HIV-induced encephalopathy, osteoclastoma and osteoporosis, and metabolic bone diseases.
In osteoclasts, TRAP is localized within the ruffled border area, the lysosomes, the Golgi cisternae and vesicles.[5]
TRAP gene, promoter organisation and transcription
Mammalian TRAP is encoded by one gene, which is localized on chromosome 19 (19p13.2-13.3) in humans, and on chromosome 9 in mice. TRAP DNA is, as expected from protein sequencing, highly conserved throughout the class mammalia. The TRAP gene has been cloned and sequenced in porcine, rat, human, and murine species.[9] Human, murine, and porcine TRAP genes all contain 5 exons, and have the ATG codon at the beginning of exon 2, with exon 1 being non-coding. Within the exon 1 promoter, there are three distinct “tissue-specific” promoters: 1A, 1B, and 1C.[10] This would allow TRAP expression to be tightly controlled. Transcribed from this gene is a 1.5kb mRNA with an open reading frame (ORF) of 969-975 bp encoding a 323-325 amino acid protein. In the rat, the ORF is 981 bp in length and encodes for a 327-amino acid protein. TRAP is translated as a single polypeptide. TRAP gene transcription is regulated by the Microphthalmia-associated transcription factor.[11][12]
Physiology
The exact physiologic role(s) of TRAP is unknown, but many functions have been attributed to this protein. In knockout studies, TRAP−/− mice exhibit mild osteopetrosis, associated with reduced osteoclast activity. These result in thickening and shortening of the cortices, formation of club-like deformities in the distal femur, and widened epiphyseal growth plates with delayed mineralization of cartilage, all of which increase with age.[13] In TRAP overexpressing transgenic mice, mild osteoporosis occurs along with increased osteoblast activity and bone synthesis.[14] Proposed functions of TRAP include osteopontin /bone sialoprotein dephosphorylation, the generation of reactive oxygen species (ROS), iron transport, and as a cell growth and differentiation factor.
Protein dephosphorylation and osteoclast migration
It has been shown that osteopontin and bone sialoprotein, bone matrix phosphoproteins, are highly efficient in vitro TRAP substrates, which bind to osteoclasts when phosphorylated.[15] Upon partial dephosphorylation, both osteopontin and bone sialoprotein are incapable of binding to osteoclasts. From this effect, it has been hypothesized that TRAP is secreted from the ruffled border, dephosphorylates osteopontin and allows osteoclast migration, and further resorption to occur.
ROS generation
Reactive oxygen species (ROS) are generated in macrophages and osteoclasts from superoxide (O2−.), which forms from the action of NADPH-oxidase on oxygen (O2).[16] They play an essential role in the function of phagocytic cells.
TRAP, containing a redox active iron, catalyzes the generation of ROS through Fenton chemistry:[17]
- O2 → (NADPH-oxidase) O2- ∙ → (superoxide dismutase) H2O2 → (catalase) H2O + O2
- TRAP-Fe3+ (purple) + O2- ∙→ TRAP-Fe2+ (pink) + O2
- H2O2 + TRAP-Fe2+ (pink) → HO∙ + HO− + TRAP-Fe3+
producing hydroxyl radicals, hydrogen peroxide, and singlet oxygen. In osteoclasts, ROS are generated at the ruffled border and seem to be required for resorption and degradation to occur.
Iron transport
In the pregnant sow, uteroferrin is highly expressed in the uterine fluids.[18] Due to the unique anatomy of the porcine uterus, and the specific, progesterone-induced expression of TRAP; it is hypothesized that uteroferrin acts as an iron transport protein.
Cell growth and differentiation factor
TRAP is associated with osteoclast migration to bone resorption sites, and, once there, TRAP is believed to initiate osteoclast differentiation, activation, and proliferation. This hypothesis was formed from the examination of the bone structure of TRAP-null mice. It was noted that, in addition to osteopetrosis, bone formation occurred in a haphazard manner, where the microarchitecture was highly irregular.[19]
In TRAP overexpressing mice, it has been found that the affected mice are grossly obese. This has led to the hypothesis that TRAP has involvement in hyperplastic obesity.
References
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- ↑ Baumbach GA, Saunders PT, Ketcham CM, Bazer FW, Roberts RM (1991). "Uteroferrin contains complex and high mannose-type oligosaccharides when synthesized in vitro". Mol. Cell. Biochem. 105 (2): 107–17. doi:10.1007/bf00227750. PMID 1922010.
- ↑ Ljusberg J, Ek-Rylander B, Andersson G (1999). "Tartrate-resistant purple acid phosphatase is synthesized as a latent proenzyme and activated by cysteine proteinases". Biochem. J. 343 (1): 63–9. doi:10.1042/0264-6021:3430063. PMC 1220524
 . PMID 10493912.
. PMID 10493912. - 1 2 Ljusberg J, Wang Y, Lång P, Norgård M, Dodds R, Hultenby K, Ek-Rylander B, Andersson G (2005). "Proteolytic excision of a repressive loop domain in tartrate-resistant acid phosphatase by cathepsin K in osteoclasts". J. Biol. Chem. 280 (31): 28370–81. doi:10.1074/jbc.M502469200. PMID 15929988.
- ↑ Klabunde T, Sträter N, Fröhlich R, Witzel H, Krebs B (1996). "Mechanism of Fe(III)-Zn(II) purple acid phosphatase based on crystal structures". J. Mol. Biol. 259 (4): 737–48. doi:10.1006/jmbi.1996.0354. PMID 8683579.
- ↑ Burstone MS (1959). "Histochemical demonstration of acid phosphatase activity in osteoclasts". J. Histochem. Cytochem. 7 (1): 39–41. doi:10.1177/7.1.39. PMID 13664936.
- ↑ Minkin C (1982). "Bone acid phosphatase: tartrate-resistant acid phosphatase as a marker of osteoclast function". Calcif. Tissue Int. 34 (3): 285–90. doi:10.1007/BF02411252. PMID 6809291.
- ↑ Cassady AI, King AG, Cross NC, Hume DA (1993). "Isolation and characterization of the genes encoding mouse and human type-5 acid phosphatase". Gene. 130 (2): 201–7. doi:10.1016/0378-1119(93)90420-8. PMID 8359686.
- ↑ Walsh NC, Cahill M, Carninci P, Kawai J, Okazaki Y, Hayashizaki Y, Hume DA, Cassady AI (2003). "Multiple tissue-specific promoters control expression of the murine tartrate-resistant acid phosphatase gene". Gene. 307: 111–23. doi:10.1016/S0378-1119(03)00449-9. PMID 12706893.
- ↑ Luchin A, Purdom G, Murphy K, Clark MY, Angel N, Cassady AI, Hume DA, Ostrowski MC (2000). "The microphthalmia transcription factor reulates expression of the tartrate-resistant acid phosphatase gene during terminal differentiation of osteoclasts". J. Bone Miner. Res. 15 (3): 451–460. doi:10.1359/jbmr.2000.15.3.451. PMID 10750559.
- ↑ Hoek KS, Schlegel NC, Eichhoff OM, Widmer DS, Praetorius C, Einarsson SO, Valgeirsdottir S, Bergsteinsdottir K, Schepsky A, Dummer R, Steingrimsson E (2008). "Novel MITF targets identified using a two-step DNA microarray strategy". Pigment Cell Melanoma Res. 21 (6): 665–76. doi:10.1111/j.1755-148X.2008.00505.x. PMID 19067971.
- ↑ Hayman AR, Jones SJ, Boyde A, Foster D, Colledge WH, Carlton MB, Evans MJ, Cox TM (1996). "Mice lacking tartrate-resistant acid phosphatase (Acp 5) have disrupted endochondral ossification and mild osteopetrosis". Development. 122 (10): 3151–62. PMID 8898228.
- ↑ Angel NZ, Walsh N, Forwood MR, Ostrowski MC, Cassady AI, Hume DA (2000). "Transgenic mice overexpressing tartrate-resistant acid phosphatase exhibit an increased rate of bone turnover". J. Bone Miner. Res. 15 (1): 103–10. doi:10.1359/jbmr.2000.15.1.103. PMID 10646119.
- ↑ Ek-Rylander B, Flores M, Wendel M, Heinegård D, Andersson G (1994). "Dephosphorylation of osteopontin and bone sialoprotein by osteoclastic tartrate-resistant acid phosphatase. Modulation of osteoclast adhesion in vitro". J. Biol. Chem. 269 (21): 14853–6. PMID 8195113.
- ↑ Darden AG, Ries WL, Wolf WC, Rodriguiz RM, Key LL (1996). "Osteoclastic superoxide production and bone resorption: stimulation and inhibition by modulators of NADPH oxidase". J. Bone Miner. Res. 11 (5): 671–5. doi:10.1002/jbmr.5650110515. PMID 9157782.
- ↑ Fenton, H.J.H., Oxidation of tartaric acid in presence of iron. J Chem Soc Trans, 1894. 65: p. 899-910.
- ↑ Roberts RM, Raub TJ, Bazer FW (1986). "Role of uteroferrin in transplacental iron transport in the pig". Fed. Proc. 45 (10): 2513–8. PMID 3527760.
- ↑ Sheu TJ, Schwarz EM, Martinez DA, O'Keefe RJ, Rosier RN, Zuscik MJ, Puzas JE (2003). "A phage display technique identifies a novel regulator of cell differentiation". J. Biol. Chem. 278 (1): 438–43. doi:10.1074/jbc.M208292200. PMID 12403789.
External links
- tartrate-resistant acid phosphatase at the US National Library of Medicine Medical Subject Headings (MeSH)
PDB gallery | ||||||
|---|---|---|---|---|---|---|
| ||||||