MGC50722
MGC50722, also known as uncharacterized protein LOC399693, is a protein that in humans is encoded by the MGC50722 gene (Mammalian Gene Collection Project Gene 50722[1]). This 965 amino acid human protein has a molecular weight of 104.495 kDa and one domain of unknown function (DUF390).[2] Generally conserved across mammals, this quickly evolving gene shows relatively low expression in most human tissues except in the testis.[3][4]
Gene
The entire human gene is 40,364 base pairs in length, while the unprocessed mRNA is 25,960 base pairs long. After splicing of introns the 10 exon gene has a final mRNA length of 3,596 base pairs that encodes for 965 amino acids.[2][5][6]
Locus
Human MGC50722 is located on the minus strand of chromosome 9 in the region q34 of the human genome (NCBI Gene ID: 399693). The most characterized gene in this region of the human genome is GPSM1, which encodes the G-protein-signaling modulator 1 protein.[7]
Homology and evolution
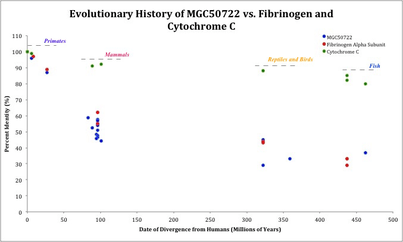
Paralogs
It was found that the centrosome-associated protein 350 (CEP350) was the only possible paralog to protein MGC50722 in humans. CEP350 is a 3117 amino acid long protein and aligns with protein MGC50722 at its N-terminus. This indicates the paralog spacing is very distant for when MGC50722 split from CEP350.
Orthologs
Compete orthologs for protein MGC50722 are found only in mammals, where most conservation is found with in the N-terminus and DUF390.
Distant homologs
The most distant homolog detectable is in cartilaginous fish (462.5 MYA).
Homologous domains
The domain of unknown function 390 (pfam04094: DUF390) is part of a family of proteins that have only been identified within the rice genome. Although this domain’s function is unknown, it may be some kind of transposable element.[8]
Protein
Primary sequence and isoforms
Human protein MGC50722 is 104.495 kDa, with an isoelectric point of 10.24. A mixed charged cluster of amino acids is present between positions 146 and 182, which seems to be conserved in primates, but not present in other mammals. There are also 6 predicted isoforms found in human.[2]
Subcellualar localization signals
PSORTII servers predict 5 nuclear localization signals in the human protein MGC50722. When ortholog sequences to the human protein were run through PSORT II, the predicted nuclear subcellular localization was a consensus prediction.
| Signal Type | Residue Span | Amino Acid Sequence |
|---|---|---|
| pat4 | 46-49 | RPRK |
| pat4 | 148-151 | KPKR |
| pat7 | 43-49 | PQQRPRK |
| pat7 | 149-155 | PKRVKSS |
| pat7 | 302-308 | PSKRRLQ |
Post-translational modifications
Human protein MGC50722 ortholog in mice, 4932418E24Rik protein, has experimentally determined phosphorylation sites at S588, S591, and S670 in the testis (pTestis ID: PT-MM-02686).[9][10][11] Prediction servers at ExPASy also predict more phosphorylation sites (NetPhos 2.0 Server), a N-termnial acetylation site (NetAcet 1.0 Server), glycation sites (NetGlycate 1.0 Server), and a GalNAc O-glycosylation site (NetOGlyc 4.0 Server) at conserved residues in the human MGC50722 protein.
Secondary structure
Prediction models characterized protein MGC50722 as mostly disordered, but two regions of coiled-coils.
Protein internal structure and features
| Feature | Residue Span |
|---|---|
| Region of Low Complexity[6] | 11-26 |
| DUF390[8][12] | 405-690 |
| Region of Low Complexity[6] | 410-423 |
| Region of Low Complexity[6] | 546-556 |
| Coiled-Coil[6] | 546-566 |
| Region of Low Complexity[6] | 606-621 |
| Coiled-Coil[6] | 720-753 |
| Region of Low Complexity[6] | 771-791 |
| Region of Low Complexity[6] | 871-884 |

Potential function
The function of protein MGC50722 is unknown. Given that it is preferentially expressed in the testis and appears to be subcellularly localized in the nucleus, it could play an important role in gamete cells.
Interacting proteins
Due the recent identification of this gene and its protein, interaction databases (MINT, STRING, IntAct, and BioGRID) have not identified any interactions. More data would expand the characterization of MGC50722.
Expression
Expression levels of human MGC50722 appear to low/absent in most cell types, with the highest and most abundant expression shown to be in the testis (GEO Profile IDs: 48997768 and 49895282).[13] A lung cancer study also showed that MGC50722 was expressed in CD4+ T-Cells of normal human tissue samples.[14]
Promoter
The transcriptional start site for MGC50722 aligns best with SPZ1, SORY, SP1F, and FAST[15] transcription factor binding sites.
Clinical significance
A significant GEO Profile relating to MGC50722 was a study done on male fertility in humans looking at the disease teratozoospermia (GEO Profile ID: 38113951).[13] Teratozoospermia is a condition where during the development of mature sperm cells morphology is altered, thus leading to, in some cases, male infertility.[16] Gene expression shows that in normal human subjects MGC50722 is expressed, while in subjects with teratozoospermia expression levels drop significantly or shut off.
References
- ↑ Strausberg RL, Feingold EA, Grouse LH, Derge JG, Klausner RD, Collins FS, Wagner L, Shenmen CM, Schuler GD, Altschul SF, Zeeberg B, Buetow KH, Schaefer CF, Bhat NK, Hopkins RF, Jordan H, Moore T, Max SI, Wang J, Hsieh F, Diatchenko L, Marusina K, Farmer AA, Rubin GM, Hong L, Stapleton M, Soares MB, Bonaldo MF, Casavant TL, Scheetz TE, Brownstein MJ, Usdin TB, Toshiyuki S, Carninci P, Prange C, Raha SS, Loquellano NA, Peters GJ, Abramson RD, Mullahy SJ, Bosak SA, McEwan PJ, McKernan KJ, Malek JA, Gunaratne PH, Richards S, Worley KC, Hale S, Garcia AM, Gay LJ, Hulyk SW, Villalon DK, Muzny DM, Sodergren EJ, Lu X, Gibbs RA, Fahey J, Helton E, Ketteman M, Madan A, Rodrigues S, Sanchez A, Whiting M, Madan A, Young AC, Shevchenko Y, Bouffard GG, Blakesley RW, Touchman JW, Green ED, Dickson MC, Rodriguez AC, Grimwood J, Schmutz J, Myers RM, Butterfield YS, Krzywinski MI, Skalska U, Smailus DE, Schnerch A, Schein JE, Jones SJ, Marra MA (Dec 2002). "Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences". Proceedings of the National Academy of Sciences of the United States of America. 99 (26): 16899–16903. doi:10.1073/pnas.242603899. PMC 139241
 . PMID 12477932.
. PMID 12477932. - 1 2 3 "Homo sapiens uncharacterized protein LOC399693". NCBI Protein.
- ↑ "Uncharacterized MGC50722 (MGC50722)". NCBI UniGene.
- ↑ Tang; et al. (2007). "Characteristics of 292 Testis-Specific Genes in Human". Biological and Pharmaceutical Bulletin. 30 (5): 865–872. doi:10.1248/bpb.30.865.
- ↑ "Homo sapiens uncharacterized MGC50722 (MGC50722), transcript variant 1, mRNA". NCBI Nucleotide.
- 1 2 3 4 5 6 7 8 9 "Transcript: MGC50722-001 ENST00000569961 Protein Summary". Ensembl.
- ↑ "MGC50722 uncharacterized MGC50722 [ Homo sapiens (human) ]". NCBI Gene. Mar 2015.
- 1 2 "Conserved Protein Domain Family DUF390".
- ↑ Qi L, Liu Z, Wang J, Cui Y, Guo Y, Zhou T, Zhou Z, Guo X, Xue Y, Sha J (Dec 2014). "Systematic analysis of the phosphoproteome and kinase-substrate networks in the mouse testis". Molecular & Cellular Proteomics. 13 (12): 3626–38. doi:10.1074/mcp.M114.039073. PMID 25293948.
- ↑ "4932418E24Rik".
- ↑ Diez-Roux G, Banfi S, Sultan M, Geffers L, Anand S, Rozado D, Magen A, Canidio E, Pagani M, Peluso I, Lin-Marq N, Koch M, Bilio M, Cantiello I, Verde R, De Masi C, Bianchi SA, Cicchini J, Perroud E, Mehmeti S, Dagand E, Schrinner S, Nürnberger A, Schmidt K, Metz K, Zwingmann C, Brieske N, Springer C, Hernandez AM, Herzog S, Grabbe F, Sieverding C, Fischer B, Schrader K, Brockmeyer M, Dettmer S, Helbig C, Alunni V, Battaini MA, Mura C, Henrichsen CN, Garcia-Lopez R, Echevarria D, Puelles E, Garcia-Calero E, Kruse S, Uhr M, Kauck C, Feng G, Milyaev N, Ong CK, Kumar L, Lam M, Semple CA, Gyenesei A, Mundlos S, Radelof U, Lehrach H, Sarmientos P, Reymond A, Davidson DR, Dollé P, Antonarakis SE, Yaspo ML, Martinez S, Baldock RA, Eichele G, Ballabio A (2011). "A high-resolution anatomical atlas of the transcriptome in the mouse embryo". PLoS Biology. 9 (1): e1000582. doi:10.1371/journal.pbio.1000582.
- ↑ "MGC50722 uncharacterized MGC50722 [Homo sapiens (human)]". NCBI Gene.
- 1 2 Barrett T, Wilhite SE, Ledoux P, Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH, Sherman PM, Holko M, Yefanov A, Lee H, Zhang N, Robertson CL, Serova N, Davis S, Soboleva A (Jan 2013). "NCBI GEO: archive for functional genomics data sets--update". Nucleic Acids Research. 41 (Database issue): D991–D995. doi:10.1093/nar/gks1193. PMC 3531084. PMID 23193258.
- ↑ Ahn, Jung-Mo; et al. (Nov 2013). "Proteogenomic Analysis of Human Chromosome 9-Encoded Genes from Human Samples and Lung Cancer Tissues". Journal of Proteome Research. 13 (1): 137–146. doi:10.1021/pr400792p.
- ↑ Yeo CY, Chen X, Whitman M (Sep 1999). "The role of FAST-1 and Smads in transcriptional regulation by activin during early Xenopus embryogenesis". The Journal of Biological Chemistry. 274 (37): 26584–90. doi:10.1074/jbc.274.37.26584. PMID 10473623.
- ↑ Machev N, Gosset P, Viville S (2005). "Chromosome abnormalities in sperm from infertile men with normal somatic karyotypes: teratozoospermia". Cytogenetic and Genome Research. 111 (3-4): 352–357. doi:10.1159/000086910.
Suggested reading
- Wagner F, French L, Veh RW (Sep 2014). "Transcriptomic-anatomic analysis of the mouse habenula uncovers a high molecular heterogeneity among neurons in the lateral complex, while gene expression in the medial complex largely obeys subnuclear boundaries". Brain Structure & Function. doi:10.1007/s00429-014-0891-9. PMID 25244943.
- Foong RE, Bosco A, Jones AC, Gout A, Gorman S, Hart PH, Zosky GR (Apr 2015). "In utero Vitamin D Deficiency Increases Airway Smooth Muscle Mass and Impairs Lung Function". American Journal of Respiratory Cell and Molecular Biology. 53: 664–75. doi:10.1165/rcmb.2014-0356OC. PMID 25867172.
- Dobrin, Radu, et al. (2009). "Multi-tissue coexpression networks reveal unexpected subnetworks associated with disease". Faculty Papers and Publications in Animal Scienced. 10: R55. doi:10.1186/gb-2009-10-5-r55. PMID 19463160.
- Ediger, BN, Du, A, Liu, J, Hunter, CS, Walp, ER, Schug, J, May, CL (Dec 2014). "Islet-1 Is essential for pancreatic β-cell function". Diabetes. 63 (12): 4206–17. doi:10.2337/db14-0096. PMID 25028525.
- Vamathevan, JJ (Jan 2009). Evolutionary analysis of mammalian genomes and associations to human disease (Thesis). University College London.