Tryptophan 2,3-dioxygenase
| TDO2 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||
| Identifiers | |||||||||||||||||
| Aliases | TDO2, TDO, TO, TPH2, TRPO, tryptophan 2,3-dioxygenase | ||||||||||||||||
| External IDs | MGI: 1928486 HomoloGene: 4132 GeneCards: TDO2 | ||||||||||||||||
| |||||||||||||||||
| RNA expression pattern | |||||||||||||||||
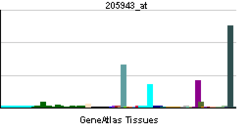 | |||||||||||||||||
| More reference expression data | |||||||||||||||||
| Orthologs | |||||||||||||||||
| Species | Human | Mouse | |||||||||||||||
| Entrez | |||||||||||||||||
| Ensembl |
|
||||||||||||||||
| UniProt | |||||||||||||||||
| RefSeq (mRNA) | |||||||||||||||||
| RefSeq (protein) | |||||||||||||||||
| Location (UCSC) | Chr 4: 155.85 – 155.92 Mb | Chr 3: 81.96 – 81.98 Mb | |||||||||||||||
| PubMed search | [1] | [2] | |||||||||||||||
| Wikidata | |||||||||||||||||
| View/Edit Human | View/Edit Mouse |
In enzymology, a tryptophan 2,3-dioxygenase (EC 1.13.11.11) is an enzyme that catalyzes the chemical reaction
- L-tryptophan + O2 N-formyl-L-kynurenine

Thus, the two substrates of this enzyme are L-tryptophan and O2, whereas its product is N-formyl-L-kynurenine. This enzyme participates in tryptophan metabolism.
In humans, tryptophan 2,3-dioxygenase is encoded by the TDO2 gene.[3]
Tryptophan 2,3-dioxygenase plays a central role in the physiological regulation of tryptophan flux in the human body. It catalyses the first and rate limiting step of tryptophan degradation along the kynurenine pathway and thereby regulates systemic tryptophan levels.[4]
Function
| tryptophan 2,3-dioxygenase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Crystal structure of the tryptophan 2,3-dioxygenase from xanthomonas campestris | |||||||||
| Identifiers | |||||||||
| EC number | 1.13.11.11 | ||||||||
| CAS number | 9014-51-1 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / EGO | ||||||||
| |||||||||
This enzyme belongs to the family of oxidoreductases, specifically those acting on single donors with O2 as oxidant and incorporation of two atoms of oxygen into the substrate (oxygenases).[5] The oxygen incorporated need not be derived from O2.[6] This family of enzymes includes tryptophan 2,3-dioxygenase (TDO, also known as tryptophan oxygenase and L-tryptophan pyrrolase) and indoleamine 2,3-dioxygenase (IDO, also known as tryptophan pyrrolase).[7] These two enzymes are oxidoreductase enzymes that contain one noncovalently bound iron–protoporphyrin IX per monomer. These enzymes catalyze the dioxygenation of L-tryptophan (L-Trp) to N-formyl-L-kynurenine in the first and rate-limiting step of the kynurenine pathway.[7]
The same family of enzymes also includes sIDO from Shewanella oneidensis[8] and PrnB, the second enzyme in the pyrrolnitrin biosynthesis pathway from Pseudomonas fluorescens,[9] although dioxygenase activity has not been demonstrated for either as yet. Recently, a new enzyme with the ability to catalyze L-tryptophan dioxygenation, INDOL1, was identified.[10]
Tryptophan 2,3-dioxygenase was initially discovered in the 1930s[11] and is found in both eukaryotes (human,[12] rat,[13] and rabbit[14]) and prokaryotes (Xanthomonas campestris,[8] and P. fluorescens[9]) Expression of tryptophan 2,3-dioxygenase in mammals is normally restricted to the liver, but it has been identified in the brain and epididymis of some species, and, in some tissues, its production can be induced in response to stimuli.[7]
Structure
Tryptophan 2,3-dioxygenase is a heme-containing cytosolic enzyme encoded by gene TDO2.[4] Crystallographic studies of xcTDO (Xanthomonas campestris TDO)[8] and rmTDO (Ralstonia metallidurans TDO)[15] have revealed that the crystal structures of xcTDO and rmTDO are essentially identical and are intimately associated homotetrameric enzymes.[16] They are best described as a dimer of dimers because the N terminal residues of each monomer form part of the substrate binding site in an adjacent monomer. The proteins are completely helical, and a flexible loop, involved in L-tryptophan binding, is observed just outside the active-site pocket. Interestingly, this loop appears to be substrate-binding induced, as it is observed only in crystals grown in the presence of L-tryptophan.[16]
The only structure available with substrate bound at the active site in the catalytically active ferrous state is xcTDO. In the structure of xcTDO, the carboxy group of L-tryptophan interacts with Arg117, Tyr113 and Thr254. Amino acid residues equivalent to Arg117 and Tyr113 are found in nearly all TDO and IDO proteins. This carboxy-binding motif appears to be essential for substrate binding; arginine reorients in the presence of substrate, co-ordinating the carboxy group of L-tryptophan. The substrate ammonium group is hydrogen-bonded to the side-chain hydroxyl group of Thr254, the 7-propionate group of the heme, and a water molecule.[16]
Mechanism
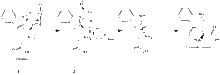
The initial formation of the ternary complex (1) occurs by substrate binding, followed by dioxygen binding to the ferrous protein. The ternary complex activates O2 and allows the otherwise spin-forbidden reaction to proceeed.[16] The formation of the hydroperoxide intermediate (2) is catalyzed by the loss of the indole proton. Two mechanisms are possible: base-catalysed deprotonation or proton abstraction by bound dioxygen.[17] However, catalysis by the ironbound dioxygen is generally proposed, as a result of experiments showing that catalytic activity is maintained upon substitution of alanine for His55 (the only basic residue in the active site of the enzyme).[8]
The rearrangement of the hydroperoxide intermediate to form the product could occur via the dioxetane intermediate (see figure) or a Criegee intermediate. However, density functional theory calculations on the catalytic mechanism of tryptophan 2,3-dioxygenase have cast doubt on the relevance of the Criegee mechanism.[18]
Clinical significance
It has been shown that tryptophan 2,3-dioxygenase is expressed in a significant proportion of human tumors.[4] In the same study, tryptophan 2,3-dioxygenase expression by tumors prevented their rejection by immunized mice. A tryptophan 2,3-dioxygenase inhibitor developed by the group restored the ability of these mice to reject tryptophan 2,3-dioxygenase-expressed tumors, demonstrating that tryptophan 2,3-dioxygenase inhibitors display potential in cancer therapy.
Another study showed that tryptophan 2,3-dioxygenase is potentially involved in the metabolic pathway responsible for anxiety-related behavior.[19] Generating mice deficient for tryptophan 2,3-dioxygenase and comparing them to the wild type, the group found that the tryptophan 2,3-dioxygenase-deficient mice showed increased plasma levels not only of tryptophan, but also of serotonin and 5-HIAA in the hippocampus and midbrain. A variety of tests, such as elevated plus maze and open-field tests showed anxiolytic modulation in these knock-out mice, the findings demonstrating a direct link between tryptophan 2,3-dioxygenase and tryptophan metabolism and anxiety-related behavior under physiological conditions.
See also
References
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- ↑ "Entrez Gene: TDO2 tryptophan 2,3-dioxygenase".
- 1 2 3 Pilotte L, Larrieu P, Stroobant V, Colau D, Dolusic E, Frédérick R, De Plaen E, Uyttenhove C, Wouters J, Masereel B, Van den Eynde BJ (Feb 2012). "Reversal of tumoral immune resistance by inhibition of tryptophan 2,3-dioxygenase". Proceedings of the National Academy of Sciences of the United States of America. 109 (7): 2497–502. doi:10.1073/pnas.1113873109. PMID 22308364.
- ↑ Ren S, Liu H, Licad E, Correia MA (Sep 1996). "Expression of rat liver tryptophan 2,3-dioxygenase in Escherichia coli: structural and functional characterization of the purified enzyme". Archives of Biochemistry and Biophysics. 333 (1): 96–102. doi:10.1006/abbi.1996.0368. PMID 8806758.
- ↑ Leeds JM, Brown PJ, McGeehan GM, Brown FK, Wiseman JS (Aug 1993). "Isotope effects and alternative substrate reactivities for tryptophan 2,3-dioxygenase". The Journal of Biological Chemistry. 268 (24): 17781–6. PMID 8349662.
- 1 2 3 Thackray SJ, Bruckmann C, Mowat CG, Forouhar F, Chapman SK, Tong L (2008). "Indoleamine 2,3-dioxygenase and tryptophan 2,3-dioxygenase". Handbook of Metalloproteins. doi:10.1002/0470028637.met223.
- 1 2 3 4 Forouhar F, Anderson JL, Mowat CG, Vorobiev SM, Hussain A, Abashidze M, Bruckmann C, Thackray SJ, Seetharaman J, Tucker T, Xiao R, Ma LC, Zhao L, Acton TB, Montelione GT, Chapman SK, Tong L (Jan 2007). "Molecular insights into substrate recognition and catalysis by tryptophan 2,3-dioxygenase". Proceedings of the National Academy of Sciences of the United States of America. 104 (2): 473–8. doi:10.1073/pnas.0610007104. PMID 17197414.
- 1 2 De Laurentis W, Khim L, Anderson JL, Adam A, Johnson KA, Phillips RS, Chapman SK, van Pee KH, Naismith JH (Oct 2007). "The second enzyme in pyrrolnitrin biosynthetic pathway is related to the heme-dependent dioxygenase superfamily". Biochemistry. 46 (43): 12393–404. doi:10.1021/bi7012189. PMID 17924666.
- ↑ Ball HJ, Sanchez-Perez A, Weiser S, Austin CJ, Astelbauer F, Miu J, McQuillan JA, Stocker R, Jermiin LS, Hunt NH (Jul 2007). "Characterization of an indoleamine 2,3-dioxygenase-like protein found in humans and mice". Gene. 396 (1): 203–13. doi:10.1016/j.gene.2007.04.010. PMID 17499941.
- ↑ Kotake Y.; Masayama I. Z. (1936). "Uber den Mechanismus der Kynureninbildung aus Tryptophan". Z. Physiol. Chem. 243: 237–44.
- ↑ Batabyal D, Yeh SR (Dec 2007). "Human tryptophan dioxygenase: a comparison to indoleamine 2,3-dioxygenase". Journal of the American Chemical Society. 129 (50): 15690–701. doi:10.1021/ja076186k. PMID 18027945.
- ↑ Allegri G, Ragazzi E, Bertazzo A, Costa CV, Rocchi R (2003). "Tryptophan metabolism along the kynurenine pathway in rats". Advances in Experimental Medicine and Biology. 527: 481–96. doi:10.1007/978-1-4615-0135-0_56. PMID 15206766.
- ↑ Allegri G, Ragazzi E, Bertazzo A, Biasiolo M, Costa CV (2003). "Tryptophan metabolism in rabbits". Advances in Experimental Medicine and Biology. 527: 473–9. doi:10.1007/978-1-4615-0135-0_55. PMID 15206765.
- ↑ Zhang Y, Kang SA, Mukherjee T, Bale S, Crane BR, Begley TP, Ealick SE (Jan 2007). "Crystal structure and mechanism of tryptophan 2,3-dioxygenase, a heme enzyme involved in tryptophan catabolism and in quinolinate biosynthesis". Biochemistry. 46 (1): 145–55. doi:10.1021/bi0620095. PMID 17198384.
- 1 2 3 4 Thackray SJ, Mowat CG, Chapman SK (Dec 2008). "Exploring the mechanism of tryptophan 2,3-dioxygenase". Biochemical Society Transactions. 36 (Pt 6): 1120–3. doi:10.1042/bst0361120. PMID 19021508.
- ↑ Sono M, Roach MP, Coulter ED, Dawson JH (Nov 1996). "Heme-Containing Oxygenases". Chemical Reviews. 96 (7): 2841–88. doi:10.1021/cr9500500. PMID 11848843.
- ↑ Chung LW, Li X, Sugimoto H, Shiro Y, Morokuma K (Sep 2008). "Density functional theory study on a missing piece in understanding of heme chemistry: the reaction mechanism for indoleamine 2,3-dioxygenase and tryptophan 2,3-dioxygenase". Journal of the American Chemical Society. 130 (37): 12299–309. doi:10.1021/ja803107w. PMID 18712870.
- ↑ Kanai M, Funakoshi H, Takahashi H, Hayakawa T, Mizuno S, Matsumoto K, Nakamura T (2009). "Tryptophan 2,3-dioxygenase is a key modulator of physiological neurogenesis and anxiety-related behavior in mice". Molecular Brain. 2 (8): 8. doi:10.1186/1756-6606-2-8. PMID 19323847.
Further reading
- Comings DE, Muhleman D, Dietz GW, Donlon T (Feb 1991). "Human tryptophan oxygenase localized to 4q31: possible implications for alcoholism and other behavioral disorders". Genomics. 9 (2): 301–8. doi:10.1016/0888-7543(91)90257-F. PMID 2004780.
- Comings DE, Muhleman D, Dietz G, Sherman M, Forest GL (Sep 1995). "Sequence of human tryptophan 2,3-dioxygenase (TDO2): presence of a glucocorticoid response-like element composed of a GTT repeat and an intronic CCCCT repeat". Genomics. 29 (2): 390–6. doi:10.1006/geno.1995.9990. PMID 8666386.
- Dick R, Murray BP, Reid MJ, Correia MA (Aug 2001). "Structure--function relationships of rat hepatic tryptophan 2,3-dioxygenase: identification of the putative heme-ligating histidine residues". Archives of Biochemistry and Biophysics. 392 (1): 71–8. doi:10.1006/abbi.2001.2420. PMID 11469796.
- Kudo Y, Boyd CA, Sargent IL, Redman CW (Mar 2003). "Decreased tryptophan catabolism by placental indoleamine 2,3-dioxygenase in preeclampsia". American Journal of Obstetrics and Gynecology. 188 (3): 719–26. doi:10.1067/mob.2003.156. PMID 12634647.
- Nabi R, Serajee FJ, Chugani DC, Zhong H, Huq AH (Feb 2004). "Association of tryptophan 2,3 dioxygenase gene polymorphism with autism". American Journal of Medical Genetics Part B. 125B (1): 63–8. doi:10.1002/ajmg.b.20147. PMID 14755447.
- Guillemin GJ, Smythe G, Takikawa O, Brew BJ (Jan 2005). "Expression of indoleamine 2,3-dioxygenase and production of quinolinic acid by human microglia, astrocytes, and neurons". Glia. 49 (1): 15–23. doi:10.1002/glia.20090. PMID 15390107.
- Tao WA, Wollscheid B, O'Brien R, Eng JK, Li XJ, Bodenmiller B, Watts JD, Hood L, Aebersold R (Aug 2005). "Quantitative phosphoproteome analysis using a dendrimer conjugation chemistry and tandem mass spectrometry". Nature Methods. 2 (8): 591–8. doi:10.1038/nmeth776. PMID 16094384.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (Oct 2005). "Towards a proteome-scale map of the human protein-protein interaction network". Nature. 437 (7062): 1173–8. doi:10.1038/nature04209. PMID 16189514.
- Baharvand H, Hashemi SM, Kazemi Ashtiani S, Farrokhi A (2006). "Differentiation of human embryonic stem cells into hepatocytes in 2D and 3D culture systems in vitro". The International Journal of Developmental Biology. 50 (7): 645–52. doi:10.1387/ijdb.052072hb. PMID 16892178.